Резюме. Кратко описывается история
изучения стрекоз (Odonata) в России. Показана роль Института систематики и
экологии животных СО РАН (Новосибирск) как центра одонатологических исследований
в стране.
Приведены основные результаты фаунистических,
зоогеографических и экологических исследований стрекоз.
Дан
список видов стрекоз России и сопредельных территорий с их распределением по
следующим географическим регионам: Европейская равнина, Кавказ, Урал, Средняя
Азия, Сибирь (включая Северный Казахстан), Российский Дальний Восток.
Abstract. Odonatological
researches in Russia.
A brief historical survey of dragonfly studies in the former USSR
is presented. The Institute of Systematics and Ecology of Animals (Russian
Academy of Sciences, Siberian Branch, Novosibirsk) has provided an important
centre for odonatological research in Russia. The basic results of faunal,
zoogeographic and ecological investigations are adduced. A list of odonate
species of Russia and adjacent territories and distribution in regions (European
plane, the Caucasus, the Ural, the Middle Asia, Siberia, the Russian Far East)
are provided.
Как известно, стрекозы не играют какой-либо заметной
роли в хозяйственной деятельности человека. Тем не менее, эти насекомые
всегда привлекали к себе внимание людей, прежде всего благодаря относительно
крупным размерам и виртуозному по лёту, что делает их очень заметными в природе
и вызывает познавательный интерес. В культуре некоторых народов, особенно в
Японии, стрекоз от носят к числу особо почитаемых и даже культовых животных.
Начало систематическому описанию таксонов стрекоз и накоплению сведений о
составе одонатофауны положил К. Линней, которому было известно 20 видов,
объединяемых им в один род. За первое столетие научной зоологии число описанных
видов возросло до 350, а родов — до 33 [Rambur, 1842]. К началу XX века
количество видов достиг ло 2220, а родов — 370 [Якобсон, Бианки, 1905].
В настоящее время науке известно около 5800 видов стрекоз, объединяемых в 886
родов [Bridges, 1993; Tsuda, 2000]. Впечатляющий рост данных о составе
одонатофауны был обеспечен участием в изучении стрекоз сотен исследователей,
внёсших выдающийся вклад в одонатологию. Так, в области фаунистики и систематики
стрекоз англичанином Ф. Фрезером (Fraser, 1880–1963) было опубликовано 280
работ, японцем С. Асахина (S. Asahina) — 215, ещё по крайней мере пять
европейских и американских учёных опубликовали более чем по 100 работ: M.
Selys-Longchamps (1813–1900), M. Lieftinck (1904–1985), N. Diasdos Santos
(1916–1989), P. Calvert (1871–1961), E. Pinhey (1910–1999). Наряду со множеством
небольших статей каждый из этих исследователей публиковал и крупные обобщения и
монографии.
В последние десятилетия интерес к изучению стрекоз
особенно возрос в связи с созданием в 1971 г. Международного общества
одонатологов. Сейчас в обществе, выделившихся из него объединениях и
национальных отделениях насчитывается не менее 2 тысяч членов, издаётся более 30
международных и национальных журналов и бюллетеней, ежегодно проводятся
симпозиумы. В наиболее крупном журнале «Odonatologica», ежеквартально издаваемом
с 1972 г., опубликовано около тысячи статей и к началу 2007 г. отреферировано
16264 одонатологических публикаций, вышедших в свет за последнюю треть века.
Существенно иначе складывалась ситуация с одонатологическими исследованиями в
России.
Несмотря на огромные размеры нашей страны, особенно в
максимальных границах Российской империи, изучением стрекоз этой территории
всегда занимались лишь единичные специалисты. В начале XX века вышла в свет
работа Г.Г. Якобсона и В.П. Бианки [1905], содержащая полную библиографию по
палеарктической одонатофауне, насчитывающую 946 работ 378 авторов. Из них только
37 публикаций принадлежали 18 авторам, непосредственно изучавшим стрекоз
Российской империи, причём подавляющее большинство этих исследований было
выполнено на территории теперешних стран Балтии, Польши, Украины и Белоруссии.
И это при том, что Россия в тех границах занимала более половины территории
Палеарктики.
В 1907 г. была опубликована первая статья
Александра Николаевича Бартенева (1882–1946), с которой начался новый период
истории отечественной одонатологии. За 40 лет плодотворной деятельности А.Н.
Бартенев опубликовал 83 работы по фауне, систематике, экологии и географическому
распространению стрекоз. Всеми остальными исследователями, вместе взятыми, за
первую половину XX века было опубликовано почти в полтора раза меньше работ по
стрекозам нашей страны (58), и этот период в российской одонатологии справедливо
называют «Бартеневским».
Вторая половина XX века в изучении
стрекоз связана прежде всего с именем Бориса Фёдоровича Белышева (1910–1993).
Период его работы в области одонатологии, как и у А.Н. Бартенева, длился сорок
лет, с 1951 г. по 1990 г., в течение которых он опубликовал 176 научных работ,
включая 7 книг.
Вся жизнь Б.Ф. Белышева прошла в Сибири. Его
жизненный путь начался в Томске, затем были Бийск, Иркутск, а с 1967 г. —
Новосибирск, где на базе Биологического института СО АН СССР (ныне Институт
систематики и экологии животных СО РАН) Б.Ф. Белышев основал уникальную по
объёму коллекцию стрекоз, добился включения одонатологических исследований в
постоянную тематику института и через аспирантуру начал создавать небольшую
группу учеников, что фактически сделало Новосибирск центром одонатологических
исследований в нашей стране. Начав с изучения одонатофауны отдельных регионов
Сибири, Б.Ф. Белышев вскоре приступил к анализу распространения стрекоз мировой
фауны, результатом которого явилась обширная серия зоогеографических публикаций,
завершившихся изданием двух написанных совместно с А.Ю. Харитоновым монографий
[1981, 1983], составивших в сумме руководство по зоогеографии стрекоз мира.
Работы А.Н. Бартенева и Б.Ф. Белышева составили золотой фонд не только
отечественной, но и мировой одонатологии и получили заслуженное международное
признание. Благодаря им было по лучено общее представление о составе
одонатофауны нашей страны и созданы оригинальные схемы зоогеографического
районирования суши на основании распространения стрекоз. Однако степень
фаунистической изученности огромной территории бывшего СССР до настоящего
времени остаётся крайне неравномерной. В лучшем положении оказались некоторые из
бывших западных союзных республик, где сложились свои национальные традиции и
группы исследователей. Наиболее изучена территория Латвии и Литвы, где долгое
время работали З.Д. Спурис [1956, 1980, 1988] и А.П. Станёните [1963, 1985].
Сравнительно хорошо изучена Украина, особенно её западная часть, обстоятельные
статьи об одонатофауне которой стали появляться ещё в XIX веке [Иванов, 1876;
Ярошевский, 1881; Родзянко, 1887, 1889]. В XX веке большой вклад в изучение
стрекоз Украины внесли Г.В. Артоболевский [1917,1927], Р.С. Павлюк [1975, 1992],
С.М. Горб [Горб и др., 2000], а в последние годы — Л.А. Хрокало [2003, 2005] и
Н.О. Матушкина [Матушкіна, Хрокало, 2002].
Относительно хорошо
изучена одонатофауна Кавказа, здесь большой вклад внёс А.Н. Бартенев [1911,
1924, 1929]. Стрекозы Закавказья известны в основном по работам Н.Н.
Акрамовского [1948].
С конца XX века одонатофауна Кавказа
изучается О.Н. Поповой [Кетенчиев, Попова, 1996; Popova, 1997], В.Ю. Стаиным
[1998], С.Г. Козьминовым [1999], Х.А. Кетенчиевым и А.Ю. Харитоновым [Кетенчиев,
Харитонов, 1998, 1999; Кетенчиев, 2002].
К хорошо изученным
территориям можно отнести Урал. Первые серии публикаций по этому региону были
сделаны А.Н. Бартеневым [1908, 1909, 1930] и Ю.М. Колосовым [1914, 1927, 1929].
С 1968 г. по настоящее время одонатофауну всей территории Урала изучает А.Ю.
Харитонов [1974, 1975, 1997], а на Южном Урале с 1992 г. работает В.А. Яныбаева
[2002, 2004; Yanybaeva et all, 2006].
На фоне сравнительно
хорошо изученной одонатофауны территорий, окружающих европейскую часть России,
сам этот регион оказался одним из наименее изученных в нашей стране. Первые
публикации по европейской России появились давно [Ульянин, 1867,1869; Полетаева,
1880; Дьяконов, 1922, 1926; Колесов, 1930а, б], но со второй половины XX века
исследования здесь почти полностью прекратились и более, чем за полстолетия, по
этому региону появилось лишь около 10 заметных публикаций [Леви, 1968; Олигер,
1970; Седых, 1974; Крыжановский, 1982].
На протяжении долгого
времени крайне слабо изученной оставалась территория Средней Азии, фауна которой
была известна по работам Ф. Брауера [1877], А.Н. Бартенева [1911, 1913], А.Н.
Поповой [1936, 1951], Б.Ф. Белышева [Белышев, Шевченко, 1958], В.Н. Крыловой
[1969, 1972]. Планомерное изучение среднеазиатской одонатофауны началось только
с середины 70 годов прошлого века [Борисов, 1985а–в; 1987; Харитонов, Борисов,
1981, 1985, 1989] и было обобщено в докторской диссертации С.Н. Борисова [2007]
по стрекозам Средней Азии.
Изучение этого региона оказалось
весьма плодотворным. Отсюда описаны единственные для нашей страны во второй
половине XX века новые для науки валидные виды стрекоз Ischnura aralensis
Haritonov, 1979 и Sympetrum haritonovi Borisov, 1983. Здесь впервые на обширном
материале удалось показать большой диапазон адаптивных стра тегий стрекоз,
объясняющий их эволюционную «живучесть», и решающую роль искусственной
оросительной системы в формировании среднеазиатской одонатофауны [Борисов,
2007].
Начало планомерному изучению одонатофауны азиатской части
России положил Б.Ф. Белышев [1951, 1952, 1953а, б, 1954, 1955а, б], который в
серии экспедиций обследовал Алтай, Нарымский край, Кулундинскую степь,
Прибайкалье, Приморье, низовья р. Лены. С 70 годов изучение сибирской фауны и
экологии стрекоз продолжается, главным образом, учениками Б.Ф. Белышева
[Харитонов, 1974, 1975, 1976а–в, 1978а, 1981, 1984а, 1986; Бе лышев, Харитонов,
1975а, 1978в; Заика, 1974, 1980, 1982]. А.Ю. Харитонов провёл экспедиционные
обследования Зауралья, юга Западной Сибири, Алтае-Саянской горной страны, юга
Приморского края, Камчатки. Им обработаны многочисленные коллекционные
материалы, поступившие за после днюю треть века в Сибирский зоологический музей
ИСиЭЖ СО РАН, и организованы многолетние стационарные работы по изучению
населения стрекоз Западно-Сибирской лесостепи на базе Чановского и Карасукского
стационаров ИСиЭЖ СО РАН.
В последние годы активный вклад в
изучение фауны стрекоз азиатской части России вносят Е.И. Маликова [1993, 1995,
1997; Haritonov, Malikova, 1998] и О.Э. Костерин [1987, 1999; Kosterin 1992,
1996, 2004, 2005; Kosterin et al., 2001]. Было проведено несколько международных
экспедиций по обследованию Камчатки, Алтая, Восточного Казахстана [Dumont et all,
2005].
В итоге этих исследований в Сибири и на Даль нем Востоке
выявлено 129 видов стрекоз. Число это довольно условно, поскольку
таксономическое положение отдельных видов нуждается в уточнении, а многие
крупные территории Сибири остаются слабо изученными или даже совсем не затронуты
ми исследованиями. В азиатской части России наиболее богата фауна стрекоз
Приморского края и Приамурья, насчитывающая 90 видов. Однако, этот регион,
особенно Среднее Приамурье, изучен ещё весьма недостаточно. Неполно выяснен
состав фауны дальневосточных островов, почти полностью отсутствуют материалы по
северо-востоку Сибири, включая Охотское побережье и огромному про странству
Средне-Сибирского плоскогорья. Мало обследованными остаются горы юга Сибири,
низо вья Оби, весь бассейн Енисея, север Сибири. Даже в наиболее изученном
регионе Сибири — в Верхнем Приобье и на Алтае — до настоящего времени случаются
неожиданные фаунистические находки.
Так, на озере Манжерок на
Алтае и в приобском Сузунском бору обнаружены локальные популяции вида
Sympetrum croceolum, считавшегося эндемичным для Дальнего Востока.
Крайне необходимы стационарные наблюдения за составом фауны и динамикой
численности от дельных видов, особенно в районах интенсивного хозяйственного
освоения и на охраняемых территориях. Нет сведений по видовому составу стрекоз
подавляющего большинства сибирских заповедников, за исключением Кедровой Пади и
Алтайского, для которых составлены относительно полные списки одонатофауны.
Неравномерность изученности одонатофауны разных регионов нашей страны достаточно
наглядно отражается в количестве публикаций по фауне стрекоз тех или иных
территорий. Можно выделить 5 категорий с разной степенью обследованности:
примерно по 25 % территории страны практически отсутствуют сведения по
одонатофауне, 35 % слабо изучены, 20 % недостаточно и только 20 % изучены
относительно хорошо. При этом за относительно хорошей изученностью стоит лишь
количество публикаций, которые не всегда образуют достаточно долгий временной
ряд, позволяющий учитывать многолетние колебания в составе фауны.
Между тем колебания эти могут быть весьма значительными, что удалось показать, в
частности, на примере одонатофауны Южного Урала [Попова, Харитонов, в печати].
Для выявления межгодовых изменений в структуре фауны стрекоз Южного Урала были
взяты, как наиболее сопоставимые, данные А.Н. Бартенева [1908] начала XX века,
данные А.Ю. Харитонова конца 60–начала 70 годов XX века и экспедиций авторов
статьи в 2004–2006 гг. — начало XXI века.
Сравнение видовых
составов выявило сильные межгодовые изменения в структуре фауны стрекоз. Так,
попарное сопоставление периодов показало, что коэффициент фаунистического
сходства П. Жаккара начала XX века и 60–70 гг. составил 54 %; 60–70 гг. и начала
XXI века — 61 %; а сравнение начала XX века с началом XXI века — 39 %. Сравнение
между тремя периодами для 25 общих видов по индексу биотической дисперсии Л.
Коха даёт величину в 43 %. Столь существенные различия в показателях
фаунистического сходства для одного и того же региона во времени сопоставимы со
значительными межрегиональными различиями, что свидетельствует о необходимости
проведения многолетнего изучения локальных и региональных фаун для получения
адекватного представления об их составе и структуре.
Полный
список видов стрекоз в пределах бывшего СССР прежде публиковался неоднократно [Белышев,
1976; Павлюк, Харитонов, 1982; Белышев и др., 1989; Спурис, 1988], но
появившиеся в последние годы новые находки, таксономические и номенклатурные
изменения потребовали очередной ревизии этого списка с уточнением распределения
таксонов по крупным географическим регионам страны (см. табл. 1).
Таблица 1. Список видов стрекоз и их
распределение по географическим регионам бывшего СССР.
Table 1.
List of dragonfly species and distribution in the regions of the former USSR.
|
Виды стрекоз |
Географические регионы |
|||||
|
Восточно-Европейская равнина |
Кавказ |
Урал |
Средняя Азия |
Сибирь |
Дальний Восток |
|
|
1 |
2 |
3 |
4 |
5 |
6 |
7 |
|
Подотряд Caloptera |
||||||
|
Calopteryx atrata Selys,1853 |
+ |
|||||
|
C. intermedia (Selys, 1887) |
+ |
|||||
|
C. japonica japonica Selys, 1869 |
+ |
+ |
||||
|
C. j. altaica Belyshev, 1955 |
+ |
|||||
|
C. orientalis (Selys, 1890) |
+1 |
|||||
|
C. samarcandica Bartenev, 1911 |
+ |
|||||
|
C. splendens splendens (Harris, 1782) |
+ |
+ |
+ |
+ |
+ |
|
|
C. transcaspica Bartenev, 1911 |
+1 |
|||||
|
C. unicolor Bartenev, 1912 |
+ |
|||||
|
C. virgo virgo (Linnaeus, 1758) |
+ |
+ |
+ |
+ |
+ |
|
|
Epallage fatime Charpentier, 1840 |
+1 |
|||||
|
Mnais pruirtosa costalis Selys, 1869 |
+ |
|||||
|
Подотряд Zygoptera |
||||||
|
Lestes barbarus (Fabricius, 1798) |
+ |
+ |
+ |
+ |
+ |
|
|
L. dryas Kirby, 1890 |
+ |
+ |
+ |
+ |
+ |
+ |
|
L. japonicus Selys, 1883 |
+ |
|||||
|
L. macrostigma (Eversmann, 1836) |
+ |
+ |
+ |
+ |
+ |
|
|
L. temporalis Selys, 1883 |
+ |
|||||
|
L. sponsa (Hansemann, 1823) |
+ |
+ |
+ |
+ |
+ |
+ |
|
L. virens virens (Charpentier, 1825) |
+ |
+ |
+ |
+ |
+ |
|
|
L. viridis parvidens Artobolevski, 1929 |
+ |
|||||
|
Sympecma fusca (Vander Linden, 1820) |
+ |
+ |
+2 |
+ |
+1 |
+ |
|
S. gobica Foerster, 1900 |
+ |
+ |
||||
|
S. paedisca (Brauer, 1877) |
+ |
+ |
+ |
+ |
+ |
+ |
|
Piatycnemis deaibata Selys et Hagen, 1850 |
+ |
+1 |
||||
|
P. pennipes pennipes (Pallas, 1771) |
+ |
+ |
+ |
+ |
+ |
+ |
|
P. phyliopoda Djakonov, 1926 |
+ |
|||||
|
Copera tokyoensis Asahina, 1948 (=Denticnemis bicolor Bartenev, 1956) |
+ |
|||||
|
Coenagrion armatum (Charpentier, 1840) |
+ |
+ |
+ |
+ |
+ |
+ |
|
C. ecornutum (Selys, 1872) |
+ |
+ |
+ |
|||
|
C. glaciale glaciale (Selys, 1872) (=C. tugur Bartenev, 1956) |
+ |
|||||
|
C. g. orientale (Belyshev, 1973) |
+ |
|||||
|
C. hastulatum (Charpentier, 1825) |
+ |
+ |
+ |
+ |
+ |
|
|
C. hylas hylas (Trybom, 1889) |
+3 |
+ |
||||
|
C. h. ussuriense (Belyshev, 1956) |
+ |
|||||
|
C. johanssoni johanssoni Wallengren, 1859 (=C. concinnum Johanssen, 1859) |
+ |
+ |
+ |
|||
|
C. j. bartenevi Belyshev, 1955 |
+ |
+ |
||||
|
С. j. corivalescens Bartenev, 1914 |
+ 1 |
|||||
|
C. lanceolatum (Selys, 1872) |
+ |
+ |
||||
|
C. lunulatum (Charpentier, 1840) (=C. vernale Hagen, 1839) |
+ |
+ |
+ |
+ |
+ |
+ |
|
C. mercuriale mercuriale Charpentier, 1840 |
+ |
|||||
|
C. ornatum Selys et Hagen, 1850 |
+1 |
+ |
+1 |
|||
|
C. puella puella (Linnaeus, 1758) |
+ |
+ |
+ |
+ |
+ |
|
|
C. pulchellum (Vander Linden, 1823) |
+ |
+ |
+ |
+ |
+ |
|
|
C. scitulum (Rambur, 1842) |
+ |
+ |
||||
|
Paracercion calamorum (Ris, 1916)* |
+ |
|||||
|
P. hieroglyphicum (Brauer, 1865)* |
+ |
|||||
|
P. plagiosum (Needham, 1930)* |
+ |
|||||
|
P. v-nigrum (Needham, 1930)* |
+ |
|||||
|
Erythromma humerale Selys, 1887 |
+1 |
+ |
||||
|
E. najas najas (Hansemann, 1823) |
+ |
+ |
+ |
+ |
+ |
|
|
E. n. baicalensis Belyshev, 1964 |
+ |
|||||
|
Erythromma viridulum viridulum Charpentier,1840 |
+ |
+ |
+9 |
|||
|
E. v. orientale Schmidt, 1960 |
+ |
|||||
|
E. lindeni Selys, 1840* |
+ |
|||||
|
Enallagma cyathigerum (Charpent.,1840) |
+ |
+ |
+ |
+ |
+ |
+ |
|
E. boreale circulatum (Selys, 1883) |
+8 |
|||||
|
Ischnura aralensis Haritonov, 1979 |
+ |
+ |
||||
|
I. asiatica (Brauer, 1865) (=I. lobata Needham, 1930) |
+ |
|||||
|
I. elegans elegans (Vander Linden, 1823) |
+ |
+ |
+ |
+ |
+ |
+ |
|
I. evansi Morton, 1919 |
+ |
|||||
|
I. forcipata Morton, 1907 |
+ |
|||||
|
I. fountainei Morton, 1905 |
+ |
+ |
||||
|
I. intermedia Dumont, 1974 |
+ 1 |
|||||
|
I. pumilio (Charpentier, 1825) |
+ |
+ |
+ |
+ |
+ 1,9 |
|
|
Nehalennia speciosa (Charpentier, 1840) |
+ |
+ |
+ |
+ |
||
|
Pyrrhosoma nymphula Sulzer, 1776 |
+ |
+ |
+ 12 |
|||
|
Подотряд Anisoptera |
||||||
|
Anisogomphus maacki (Selys, 1872) |
+ |
|||||
|
Anormogomphus kiritshenkoi Bartenev, 1913 |
+ |
|||||
|
Asiagomphus melanopsoides (Doi, 1943) |
+ |
|||||
|
Davidius lunatus (Bartenev, 1914) |
+ |
|||||
|
Gomphidia confluens Selys, 1878 |
+ |
|||||
|
Gomphus schmidti Asahina, 1956 |
+ |
|||||
|
G. vulgatissimus vulgatissimus (Linnaeus, 1758) |
+ |
+ |
+ |
|||
|
G. v.schneiderii Selys, 1850 |
+ |
+ 1 |
||||
|
Lindenia tetraphylla (Vander Linden, 1825; |
+ |
+ |
||||
|
Nihonogomphus ruptus (Selys, 1858) |
+ |
+ |
||||
|
Onychogomphus assimilis assimilis (Schneider, 1845) |
+ |
+ 1 |
||||
|
O. a. fulvipennis (Bartenev, 1912) |
+ |
|||||
|
O. flexuosus (Schneider, 1845) |
+ |
+ |
||||
|
O. forcipatus forcipatus (Linnaeus, 1758) |
+ |
+ |
+ |
|||
|
O. f. albotibialis Schmidt, 1954 |
+ 1 |
|||||
|
O. f. unguiculatus (Vander Linden, 1820J |
+ |
|||||
|
O. lefebvrei (Rambur, 1842) |
+ |
+ |
||||
|
Ophiogomphus cecilia Fourcroy, 1785 |
+ |
+ |
+ |
+ |
||
|
O. obscurus Bartenev, 1909 |
+ |
+ |
||||
|
O. reductus Calvert, 1898 |
+ |
+ 1 |
||||
|
O. spinicornis Selys, 1878 |
+ 1 |
|||||
|
Shaogomphus postocularis epophthalmus Selys, 1872 |
+ 1 |
+ |
||||
|
Sieboldius albardae Selys, 1886 |
+ |
|||||
|
Sinictinogomphus clavatus clavatus (Fabricius, 1775) |
+ |
|||||
|
Stylurus annulatus (Djakonov, 1926) |
+ |
|||||
|
S. flavipes flavipes (Charpentier, 1825) |
+ |
+ |
+ |
+ |
+ |
|
|
S. f. liniatus (Bartenev, 1929) |
+ |
+ |
||||
|
S. f. sibirica (Bartenev, 1910) |
+ |
+ |
||||
|
Trigomphus anormolobatus Bartenev, 1911 |
+ |
|||||
|
T. citimus citimus (Needham, 1931) |
+ |
|||||
|
T. nigripes (Selys, 1887) |
+ |
|||||
|
Anotogaster sieboldii (Selys, 1854) |
+8 |
|||||
|
Cordulegaster boltonii boltonii (Donovan, 1807) |
+ |
+2 |
||||
|
C. insignis charpentieri (Kolenati, 1846) |
+ |
+ 11 |
||||
|
C. i. nobilis Morton, 1916 |
+ |
|||||
|
C. i. coronatus (Morton, 1916) |
+ |
|||||
|
Cordulegaster pictus Selys, 1854 |
+ |
|||||
|
C. mzymtae Bartenev, 1930 |
+ |
|||||
|
Anax imperator imperator Leach, 1815 |
+ |
+ |
+ |
+ |
||
|
A. junius Drury, 1770 |
+7 |
|||||
|
A. parthenope parthenope Selys, 1839 |
+ |
+ |
+ |
+ |
+ 1 |
|
|
A. p. julius Brauer, 1865 |
+ |
|||||
|
Hemianax ephippiger (Burmeister, 1839) |
+ |
+ |
+ |
|||
|
Aeshna affinis Vander Linden, 1820 |
+ |
+ |
+ |
+ |
+ |
|
|
Ae. caerulea (Strom, 1783) |
+ |
+ |
+ |
+ |
+ |
|
|
Ae. crenata crenata Hagen, 1856 |
+ |
+ |
+ |
+ |
+ |
|
|
Ae. c. nigroflava Martin, 1908 |
+ |
|||||
|
Ae. cyanea (Mueller, 1764) |
+ |
+ |
+ |
|||
|
Ae. grandis (Linnaeus, 1758) |
+ |
+ |
+ |
+ |
+ |
|
|
Ae. juncea (Linnaeus, 1758) |
+ |
+ |
+ |
+ |
+ |
+ |
|
Ae. mixta mixta Latreille, 1805 |
+ |
+ |
+ |
+ |
+ |
+ |
|
Ae. serrata Hagen, 1856 |
+ |
+ |
+ |
+ |
+ |
+ |
|
Ae. subarctica subarctica Walker, 1908 |
+ |
+ |
+ |
+ |
||
|
Ae. viridis Eversmann, 1836 |
+ |
+ |
+ |
+ |
+ |
|
|
Aeschnophlebia kolthoffi Sjostedt, 1925 |
+ |
|||||
|
Ae. longistigma Selys, 1883 |
+ |
|||||
|
Anaciaeschna isoceles antehumeralis Schmidt, 1950 |
+ |
+ |
+ |
+ |
||
|
Brachytron pratense Mueller, 1764 |
+ |
+ |
+ |
|||
|
Caliaeschna microstigma (Schneider, 1845J |
+ |
+ |
||||
|
Cordulia aenea aenea (Linnaeus, 1758) |
+ |
+ |
+ |
+ |
+ |
|
|
C. a. amurensis Selys, 1887 |
+ |
+ |
||||
|
Epitheca bimaculata bimaculata (Charpentier, 1825) |
+ |
+ |
+ |
|||
|
E. b. altaica Belyshev, 1951 |
+ |
|||||
|
E. b. sibirica Selys, 1887 |
+ |
|||||
|
Epophthalmia elegans (Brauer, 1865) |
+ |
|||||
|
Macromia amphigena fraenata Martin, 1907 |
+6 |
+ |
+ |
|||
|
M. manchurica Asahina, 1964 |
+ 1 |
|||||
|
Somatochlora alpestris (Selys, 1840) |
+ |
+ |
+ |
+ |
+ |
|
|
S. arctica (Zetterstedt, 1840) |
+ |
+ |
+ |
+ |
+ |
+ |
|
S. exuberata Bartenev, 1910 |
+ |
+ |
||||
|
S. flavomaculata (Vander Linden, 1825) |
+ |
+ |
+ |
+ |
||
|
Somatochlora graeseri graeseri Selys, 1887 |
+ |
+ |
+ |
|||
|
S. g. aureola Oguma, 1915 |
+ |
|||||
|
S. metallica metallica (Vander Linden, 1825) |
+ |
+ |
+ |
+ |
||
|
S. m. abacanica Belyshev, 1955 |
+ |
|||||
|
S. sahlbergi Trybom, 1889 |
+3 |
+ |
+ |
|||
|
S. uchidai Foerster, 1909 |
+ |
|||||
|
S. viridiaenea (Uhler, 1858) |
+ |
|||||
|
Crocothemis erythraea chaldaeorum Morton, 1920 |
+ |
+ |
||||
|
C. servilia (Drury, 1770) |
? |
+ |
||||
|
Deielia phaon (Selys, 1883) |
+ |
|||||
|
Diplacodes lefebvrei (Rambur, 1842) |
+ 1 |
|||||
|
Leucorrhinia albifrons (Burmeister, 1839) |
+ |
+ |
+ |
|||
|
L. caudalis (Charpentier, 1840) |
+ |
+ |
+ |
+ |
||
|
L. circassica Bartenev, 1929 |
+ |
|||||
|
L. dubia (Vander Linden, 1825) |
+ |
+ |
+ |
+ |
||
|
L. intermedia Bartenev, 1910 |
+ |
+ |
||||
|
L. orientalis Selys, 1887 (=L. ussuriensis Bartenev, 1914) |
+3 |
+ |
+ |
|||
|
L. pectoralis (Charpentier, 1825) |
+ |
+ |
+ |
+ |
+ |
|
|
L. rubicunda (Linnaeus, 1758) |
+ |
+ |
+ |
|||
|
Libellula depressa Linnaeus, 1758 |
+ |
+ |
+ |
+ |
+ 1 |
|
|
L. fulva fulva Mueller, 1764 |
+ |
+ |
+ 12 |
|||
|
L. f. pontica Selys, 1887 |
+ |
|||||
|
L. quadrimaculata quadrimaculata Linnaeus, 1758 |
+ |
+ |
+ |
+ |
+ |
|
|
L. q. orientalis Belyshev, 1956 |
+ |
|||||
|
Lyriothemis pachygastra (Selys, 1878) |
+ |
|||||
|
Neurothemis fluctuans (Fabricius, 1793) (=N. palliata palliata (Rambur, 1842)) |
+ |
|||||
|
Orthetrum albistylum albistylum (Selys, 1848) |
+ |
+ |
+ 1 |
|||
|
O. a. speciosum (Uhler, 1858) |
+ |
|||||
|
Orthetrum anceps (Schneider, 1845) |
+1 |
+ |
+ |
|||
|
O. brunneum brunneum Fonscolombe, (1837) |
+1 |
+ |
+ |
+5 |
||
|
O. cancellatum cancellatum (Linnaeus, 1758) |
+ |
+ |
+ |
|||
|
O. c. kraepelini Ris,1897 |
+ 1 |
|||||
|
O. c. orientalis Belyshev, 1958 |
+ |
|||||
|
O. coerulescens Fabricius,1798 |
+ |
|||||
|
O. melania (Selys, 1883) (=O. triangulare (Selys, 1878)) |
+8 |
|||||
|
O. sabina (Drury, 1770) |
+ |
+ |
||||
|
Pantala flavescens (Fabricius, 1798) |
+ |
+ |
+ |
|||
|
Selysiothemis nigra (Vander Linden, 1825) |
+ |
+6 |
+ |
|||
|
Sympetrum arenicolor Jodicke, 1994 (=S. vulgatum flavum Bartenev, 1915)** |
+ |
+ |
||||
|
S. baccha matutinum Ris, 1911 |
+ |
|||||
|
S. cordulegaster (Selys, 1883) |
+ |
|||||
|
S. croceolum (Selys, 1883) |
+9 |
+ |
||||
|
S. danae (Sulzer, 1776) |
+ |
+ |
+ |
+ |
+ |
+ |
|
S. depressiusculum (Selys, 1841) |
+ |
+ |
+ |
+ |
||
|
S. eroticum eroticum (Selys, 1883) |
+ 10 |
+ |
||||
|
S. flaveolum flaveolum (Linnaeus, 1758) |
+ |
+ |
+ |
+ |
+ |
+ |
|
S. fonscolombii (Selys, 1840) |
+ |
+ |
+ |
|||
|
S. frequens (Selys, 1883) |
+ |
|||||
|
S. haritonovi Borisov, 1983 |
+ |
|||||
|
S. infuscatum (Selys, 1883) |
+ |
|||||
|
S. imitans (Selys, 1886) |
+ |
|||||
|
S. kunckeli (Selys, 1884) |
+ |
|||||
|
S. meridionale (Selys, 1841) |
+ |
+ |
+ |
+9 |
||
|
S. parvulum (Bartenev, 1912) |
+ |
|||||
|
S. pedemontanum pedemontanum (Mueller, 1766) |
+ |
+ |
+ |
+ |
+ |
+ |
|
S. p. elatum (Selys, 1872) |
+8 |
|||||
|
S. risi Bartenev, 1914 |
+ |
|||||
|
S. sanguineum sanguineum (Mueller, 1764) |
+ |
+ |
+ |
+ |
+ |
|
|
S. s. armeniacum Selys, 1884 |
+ |
|||||
|
S. striolatum striolatum (Charpentier, 1840) |
+ |
+ |
+ |
+4 |
||
|
S. s. imitoides Bartenev, 1919 |
+ |
|||||
|
S. s. kurilis Belyshev, 1976 |
+8 |
|||||
|
S. s. pallidum (Selys, 1887) |
+ |
|||||
|
S. tibiale Ris, 1897 |
+ |
+ |
||||
|
S. vulgatum vulgatum (Linnaeus, 1758) |
+ |
+ |
+ |
|||
|
S. v. decoloratum (Selys, 1884) |
+ |
+ |
||||
|
S. uniforme (Selys, 1883) |
+ |
|||||
|
Trithemis festiva (Rambur, 1842) |
+ 1 |
|||||
|
Всего видов: 172 |
72 |
85 |
70 |
84 |
75 |
94 |
* — название и систематическое положение таксона дано по [Weekers, Dumont,
2004]; ** — название вида дано по [Jodicke et all., 2000]. 1 — юг региона; 2 —
данные Боев и др., 1989 для Башкирии; 3 — только Полярный Урал; 4 — только
Юго-Западный Алтай; 5 — данные Hagen [1856]; 6 — единственная находка авторов
статьи на Южном Урале; 7 — данные Hagen [1856] для Камчатки; 8 —
только дальневосточные острова; 9 — крайне локально; 10 — единственная находка в
Литве [Станёните, in lit.]; 11 — только Юго-Западный Копетдаг; 12 — данные
Зей-Нечаевой, Баянова [1975] для Башкирии.
* — taxonomy is given
by Weekers, Dumont [2004]; ** — species name is given according to Jodicke et
all. [2000]. 1 –South of region; 2 — according to Boev et al. [Боев и др., 1989]
for Bashkiria; 3 — Polar Urals only; 4 — South-West Altai only; 5 — from Hagen
[1856]; 6 — unique finding in Southern Urals; 7 — from Hagen [1856] for
Kamtchatka; 8 — only Far Eastern islands; 9 — strictly local; 10 — unique
finding in Litva [Stanenite, in lit.]; 11 — South-West Kopetdagh only; 12 — from
Zei-Nechaeva and Bayanov [ЗейНечаева, Баянов, 1975] for Bashkiria.
Из публиковавшихся прежде списков одонатофауны выведен
ряд таксонов, которые в последние годы были сведены в синонимы. В соответствии с
ревизией ряда дальневосточных таксонов стрекоз [Маликова, 1995] вид
Aeschnophlebia zygoptera Belyshev,1965 сведён в синоним Aeschnophlebia
kolthoffi Sjostedt, 1925; вид Temnogomphus amurensis Bartenev, 1930 —
в синоним Anisogomphus maacki (Selys, 1872); вид Gomphus chankae
Bartenev, 1956 — в синоним G. schmidti Asahina, 1956; вид Coenagrion
striatum Bartenev, 1956 — в синоним Cercion plagiosum (Needham,
1930); вид Coenagrion brevicauda Bartenev, 1956 — в синоним Cercion
v-nigrum (Needham, 1930). Виды Macromia sibirica Djakonov, 1926 и
Macromia bartenevi Belyshev, 1973 сведены в синонимы Macromia amphigena
fraenata Martin, 1907 [Malikova, 1995].
Вид Gomphus
epophthalmus Selys, 1872 в соответствии с системой, принятой сейчас рядом
авторов [Kosterin, 2004], переведён в таксон Shaogomphus postocularis
epophthalmus Selys, 1872. Вид Libellula relicta Belyshev et Kiauta,
1968 сведён в синоним Libellula quadrimaculata ab. praenubila Newman [Ketenchiev,
Haritonov, 1999].
Существенные преобразования произведены с
таксонами рода Aeshna. По коллекциям ИСиЭЖ СО РАН, других российских и
некоторых европейских и американских музеев, а также специальным сборам в
природе была оценена морфологическая изменчивость более 3000 экземпляров имаго,
3500 экзувиев и 2000 личинок всех палеарктических и др.
представителей рода Aeshna [Белевич, 2005]. В результате было
установлено, что самый широкораспространённый вид рода, Ae. juncea,
обладает большой индивидуальной изменчивостью как окрасочных, так и структурных
признаков, что при вело к необоснованному описанию в Палеарктике шести его
подвидов. Признано целесообразным все эти подвиды, а также виды Aeshna
undulata Bartenev, 1930 и Ae. baicalensis Belyshev, 1964 свести в
синонимы Ae. juncea. Было показано, что описанный ранее подвид Aeshna
crenata wnukovskii Belyshev, 1973 также не обоснован и должен быть сведён в
синоним номинативной формы, а вид Aeshna nigroflava Martin, 1908 следует
считать подвидом Aeshna crenata, населяющим Японские и другие дальне
восточные острова. Подвид Aeshna serrata osiliensis Mierzejwski, 1913
является синонимом номинативной формы Aeshna serrata. Вид Aeshna
palmata Hagen, 1856 также выводится из состава азиатской одонатофауны, так
как указание этого североамериканского вида для Камчатки явно явилось
результатом ошибки с географическими этикетками [Gloyd, 1941].
Существует много противоречивых мнений относительно палеарктических таксонов
рода Enallagma. Для решения этой проблемы было пред принято специальное
исследование, в ходе которого были сравнены серии признаков почти у 1500
экземпляров представителей рода [Юрченко, 2004].
В результате
было сделано заключение, что всю материковую часть Палеарктики населяет
единственный вид Enallagma cyathigerum Charpentier, 1840, сильная
морфологическая изменчивость которого привела к ошибочному описанию ряда видов и
подвидов. По мнению этого автора, которое представляется достаточно
обоснованным, описанные ранее виды Enallagma deserti Selys, 1871; E.
risi Schmidt, 1961; E. strouhali Quentin, 1962; E. nigrolineata
Belyshev et Haritonov, 1975 целесообразно свести в синонимы полиморфного вида
E. сyathigerum.
Возможно лишь, что E. risi может быть
наделён статусом подвида Enallagma cyathigerum risi Schmidt, 1961,
поскольку существует клинальная изменчивость признаков от северных к южным
популяциям. Однако если наиболее северные и наиболее южные популяции хорошо
дифференцируются как подвиды E. c. cyathigerum и E. c. risi, то на
огромной промежуточной территории, занимающей почти всю умеренную зону России,
признаки подвидов перемешиваются и их идентификация становится затруднительной
или невозможной. Таксон E. circulatum Selys, 1883, вероятно, следует
считать островным азиатским подвидом североамериканского вида E. boreale
Selys, 1875, а описанный ранее вид E. belyshevi Haritonov, 1975
целесообразно свести в синоним подвида E. boreale circulatum.
Изучение больших серий палеарктических видов рода Sympetrum [Haritonov,
Popova, 1997; Попова, 1998, 1999; Popova, 2002, 2004] выявило у некоторых из них
сильную модификационную, в частности, клинальную изменчивость. В результате у
вида S. pedemontanum сведены в синонимы его подвиды S. p. intermedium
Belyshev, 1956 и S. p.kurentzovi Belyshev, 1956 и признано, что у этого
вида имеется только два подвида — номинативный на материке и подвид S. p.
elatum Selys, 1883 на дальневосточных островах. Сведены в синонимы к
номинативным формам подвиды Sympetrum striolatum dochidordzi Belyshev,
1958 и S.croceolum fuscoatrum Belyshev, 1964. Описанные Б.Ф. Белышевым
подвиды Somatochlora flavomaculata minor, S. sahlbergi relicta,
Leucorrhinia albifrons obensis сводятся в синонимы к номинативным формам,
так как были описаны на недостаточном материале без учёта индивидуальной
изменчивости и их подвидовой статус впоследствии не подтвердился.
Таким образом, в представленном в данной статье уточнённом списке стрекоз
бывшего СССР насчитывается 172 вида, 25 из которых имеют на этой территории по
2–3 подвида. Шесть приведённых в таблице регионов различаются по размерам,
природно-климатическим условиям, и специфике одонатофауны. Наиболее богата и
специфична преимущественно автохтонная фауна стрекоз южной части Дальнего
Востока. Это самая древняя и самобытная для азиатской части России фауна,
благодаря чему она выделяется в особую Субголарктическую фаунистическую область.
Остальная территория входит в состав Голарктической фаунистической области и
разделяется на две подобласти: Европейско-Сибирскую (Урал и Западная Сибирь) и
Сибирскую (Восточная Сибирь и север Дальнего Востока). Фауна Голарктической
области обеднена, исторически молода и аллохтонна, то есть сформировалась в
послеледниковое время за счёт миграций с соседних территорий.
Как уже отмечалось, наряду с фаунистическими исследованиями в отечественной
одонатологии сложились давние зоогеографические традиции.
Если в
зарубежной литературе почти отсутствует ареалогический анализ и попытки
зоогеографического районирования на основании распространения стрекоз, то в
отечественной литературе уже А.Н. Бартеневым [1933а, б; 1932а, 1935, 1939] были
сделаны оригинальные и широкие обобщения на одона тологическом материале.
Выдающийся вклад в описание и интерпретацию географического распространения
стрекоз внёс Б.Ф. Белышев, более половины публикаций которого посвящены этим
проблемам [Белышев 1956, 1959, 1960а]. Эти традиции продолжены А.Ю. Харитоновым
[1972, 1978б, 1991] и в большой серии их совместных работ с Б.Ф. Белышевым [Белышев,
Харитонов, 1975б, 1978, 1980, 1981, 1983], С.Н. Борисовым [1987, 2002, 2007],
О.Н. Поповой [1999, 2001а, б], Х.А. Кетенчиевым [2000, 2002а, б].
Много работ в мировой литературе посвящено поведению стрекоз. Проводятся такие
исследования и в нашей стране. В частности, оригинальные экспериментальные
работы по поведению стрекоз, связанному с их топическими и субстратными
предпочтениями, провёл В.В. Заика [1977, 1982]. Одна ко в отличие от зарубежных
работ большее внимание в нашей стране уделяется поведению стрекоз на личиночной
фазе развития. Эту традицию заложила сотрудница кафедры энтомологии МГУ Г.И.
Рязанова. Результаты её интересных и изящных экспериментов отражены в большой
серии публикаций [Рязанова, Мазохин-Поршняков, 1984, 1985, 1989] и обобщены в
докторской диссертации [1998]. В Сибири работы по поведению личинок стрекоз
продолжены М.В. Дронзиковой [1994, 2000, 2004].
Сильная
российская паразитологическая школа также вовлекла стрекоз в сферу исследований
про межуточных хозяев в жизненных циклах гельминтов и других паразитов. На эту
тему выполнен цикл работ, в том числе кандидатских диссертаций, по разным
регионам: Латвии [Станёните, 1963], Украине [Павлюк, 1972, 1975], Южному Уралу
[Баянов, 1974; Гуляев, 1989; Ткачёв, 1969, 1970], Казахстану [Кукашев, 1985,
1989], югу Западной Сибири [Илюшина, 1973, 1975; Карпенко, Заика, 1979; Крюкова,
2005; Сербина, 2002].
Однако наиболее ярко развивающееся
направление отечественной одонатологии — экология стрекоз. Основы экологических
подходов в изучении стрекоз были заложены в 30 годах прошлого века в работе В.Г.
Колесова [1930б] «Экология Odonata Московской губернии» и в больших статьях А.Н.
Бартенева [1930, 1932б] о биологических группировках видов стрекоз. Ценные
данные по экологии стрекоз приведены в некоторых статьях Б.Ф. Белышева [1960б,
1962, 1964, 1966] и его книге «Стрекозы Сибири» [1973, 1974].
Экология стрекоз стала основным направлением исследований А.Ю. Харитонова —
ученика и преемника Б.Ф. Белышева. В его докторской диссертации «Бореальная
одонатофауна и экологические факторы географического распространения стрекоз»
[1991а] впервые был выявлен широкий набор этолого-климатических адаптаций
стрекоз и показано, что одним из основных факторов в их распространении служат
межвидовые взаимодействия.
Конкуренция за наиболее
лимитированный ресурс — места яйцекладок — приводит к насыщению ёмкости
репродуктивных стаций ограниченным числом видов и становлению устойчивого по
составу и структуре населения, препятствующего вселению новых видов, что во
многом объясняет феномен относительной стабильности границ ареалов у этих высоко
подвижных и экологически пластичных насекомых.
Этот подход к
изучению стрекоз был продолжен и развит в работах С.Н. Борисова [1985а–в; 1987,
2006а–в] и обобщён им в докторской диссертации «Стрекозы Средней Азии и их
адаптивные стратегии» [2007]. В частности, им было выяснено, что особи ряда
среднеазиатских видов из равнинных популяций летом сразу после выплода улета ют
в горы, а осенью возвращаются на равнины для размножения или зимовки имаго, что
позволяет им избегать воздействия экстремально высоких летних температур и
сухости на равнинах. Для некоторых широко распространённых видов были выявлены
сезонные широтные миграции: в ранневесенний период стрекозы прилетают из
тропических частей ареалов на территорию Средней Азии, где происходит развитие
их летних генераций во временных водоёмах с несложившимися гидробиоценозами, что
в значительной степени снижает сопротивление аборигенной биоты.
В серии кандидатских диссертационных исследований аспирантов А.Ю. Харитонова
были изучены эколого-фаунистические особенности отдельных региональных фаун.
Так, в диссертациях Г.А. Сухачёвой (Смирновой) [1989] впервые были количественно
оценены некоторые параметры трофических связей стрекоз в Западно-Сибирской
лесостепи; Е.И. Маликовой [1995] — описаны фенология и биотопическое
распределение стрекоз на Дальнем Востоке России; С.Г. Козьминовым [1999] и В.Ю.
Стаиным [2002] — особенности личиночного развития и высотно-поясного
распределения видов в горах Кавказа; М.В. Дронзиковой [2000] — показаны
региональные особенности в структуре населения, жизненных циклах и поведении
личинок стрекоз в бассейне реки Томи; В.А. Яныбаевой [2002] — получены
количествен ные данные по населению имаго и влиянию антро погенных загрязнений
на стрекоз Южного Урала; М.А. Бекетовым [2003] — применению стрекоз в
биомониторинге и экотоксикологических исследованиях в Верхнем Приобье; И.А.
Чаплиной [2004] — выявлены перестройки структуры населения стрекоз в Восточном
Казахстане под влиянием различных экологических факторов. Ряд диссертаций
посвящён таксономическим ревизиям отдельных родов стрекоз, а также экологии
входящих в них видов и соответствующих таксоценов в Палеарктике. В этот ряд
входят диссертационные исследования О.Н. Поповой [1999] по роду Sympetrum,
Ю.А. Юрченко [2004] по роду Enallagma, О.Э. Белевич [2005] по роду
Aeschna.
В последние годы особое внимание уделяется изучению
населения личинок и имаго стрекоз как единого целого с перспективой
количественной оценки вклада этих насекомых в трофические и паразитарные циклы,
круговорот вещества и энергии и другие биогеоценотические процессы. Получены
первые оценки места и роли стрекоз в биоценозах западно-сибирской лесостепи. Для
этого ландшафта масса личинок стрекоз в водоёмах может достигать 130 г/м2
(в сыром весе) и составлять половину и даже более общей зоомассы фитофильных
сообществ; суммарная за сезон плотность имаго в оптимальных местообитаниях
превышает 0,5 млн. особей, а биомасса — 40 и более кг/га [Харитонов, 1991].
Абсолютное большинство работ с количественными оценками популяций или населения
стрекоз выполнено раздельно по личинкам или имаго этих насекомых. В настоящее
время авторам статьи удалось преодолеть ряд методических трудностей и проводить
комплексные оценки населения как водной, так и наземной фаз развития [Попова,
2001в, 2004; Попова, Харитонов, 1996, 1998, 2004; Харитонов, Попова, 1993], что
позволило получить новые интересные результаты. Например, в отношении стрекоз
общеизвестно, что сразу после выплода молодые имаго с водоёма отлетают в
околоводные биотопы, где они кормятся, находят укрытие от хищников и непогоды,
ночуют и по достижении половой зрелости начинают возвращаться на водоём, чтобы
оста вить здесь потомство. Однако на барабинских водоёмах с тростниковыми
зарослями бордюрного или займищного типа может происходить внутри популяционная
дифференциация у некоторых видов стрекоз, при которой возникают разные
топические группировки — плёсовые, не покидающие водоём, и тростниковые,
отлетающие с водоёма на берег по «стандартной» схеме [Попова, 2006].
Возрастает спрос на работы по стрекозам как одному из компонентов
биоразнообразия, связанный, в частности, с составлением региональных каталогов
животного мира, баз данных, созданием особо охраняемых природных территорий и
Красных книг [Харитонов, 1984б, 1987, 1989, 2000, 2001; Харитонов, Попова, 1997;
Haritonov, 2001; Kalkman et al., 2004; Kosterin et al., 2004].
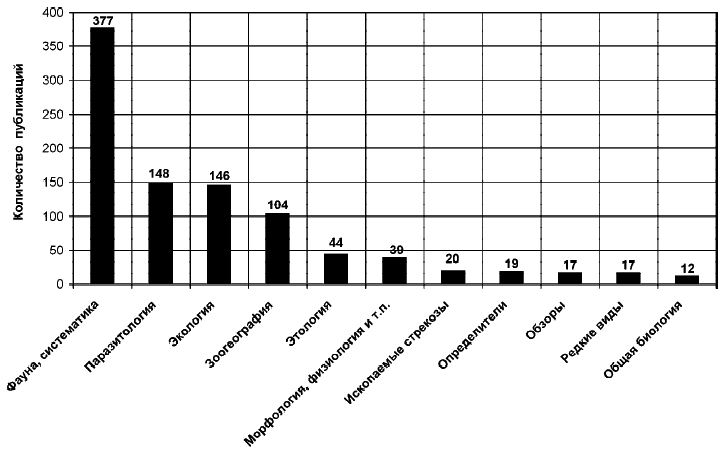
Степень разработанности каждого из перечисленных выше направлений исследований
наглядно показывает количество публикаций (рис. 1).
Рис. 1. Тематическая структура публикаций по стрекозам бывшего
СССР (общее число 943).
Fig. 1. Thematic structure of
publications on odonates of the former USSR (943 publications totally)
Сложившаяся в Институте систематики и экологии
животных СО РАН одонатологическая школа обеспечивает развитие всех этих
направлений.
Вместе с тем, существует ряд проблем, решение
которых могло бы активизировать дальнейшие работы и дополнительно привлечь к ним
молодых исследователей. Прежде всего, это отсутствие общей сводки по стрекозам
России, содержащей повидовые очерки с описанием внутривидовой система тики и
аутэкологии видов. Капитальный двухтомный труд Б.Ф. Белышева «Стрекозы Сибири»
[1973, 1974] лишь частично восполняет этот пробел, так как описывает только
сибирских стрекоз, а многие сведения из этого труда требуют существенного
дополнения и корректировки в свете новых данных, полученных в последние
десятилетия большим коллективом исследователей. Остро ощущается нехватка
определителей стрекоз — как общероссийского, так региональных. Определительные
таблицы стрекоз в академических изданиях по европейской части [Спурис,
1964] и Дальнему Востоку СССР [Харитонов, 1986] очень лаконичны и в
определённой мере устарели, по таким крупным регионам со своеобразной фауной,
как Средняя Азия и Урал, их пока нет совсем. Остаются некоторые методические
трудности в изучении миграций стрекоз, биотопического и микростациального
распределения личинок, особенно в колоссальных по площади тростниковых зарослях,
представляющих, по-видимому, одно из основных личиночных местообитаний, по
крайней мере на юге Западной Сибири и в Казахстане. У многих видов слабо
изучена изменчивость в пределах их ареалов, что не позволяет разработать
обоснованную внутривидовую систематику. При ведённые выше примеры с ревизиями
палеарктических представителей родов Aeshna, Enallagma и
Sympetrum, в результате которых было сведено в синонимы полтора десятка
необоснованных видов и подвидов, наглядно показывают актуальность и
эффективность такой работы. Практически совсем не изучена генотипическая,
модификационная и географическая изменчивость личинок стрекоз, хотя есть
основания полагать, что размах этой изменчивости может быть шире, чем у имаго.
Однако сложившиеся в отечественной одонатологии прочные традиции и
подготовленные кадры позволяют надеяться на хорошие перспективы в разностороннем
изучении стрекоз и использовании их как модельных объектов во многих
биологических исследованиях.
| © Стрекозы
Южного Урала, 2009-2010 © REDBOOK.RU, 2009-2010 |
|